La evolución de los primates
Compartir
Datos principales
Rango
Prehistoria
Desarrollo
No existe unanimidad en la taxonomía (clasificación biológica con criterios jerarquizados) de los primates entre todos los investigadores, aunque las discrepancias mayores proceden de la posición de los especímenes fósiles. Ambos criterios, clasificación y significado evolutivo, están relacionados ya que colocar un grupo de fósiles en casilleros diferentes implica, al menos, la separación en el pasado de una población ancestral en varias distintas (proceso de especiación). Estos mecanismos evolutivos, aunque siguen siendo objeto de discusión, están implícitos en cualquier clasificación taxonómica. Así, por ejemplo, si se acepta que la superfamilia de los Hominoideos se subdivide en dos familias (Hominidae e Hilobatidae) en vez de en tres, como era convencional hasta hace poco (Hominidae, Hilobatidae y Pongidae) es porque se considera relevante la temprana separación genética de los gibones, descubierta en el registro fósil, respecto a los demás hominoideos (hombres y póngidos) cuya diferenciación fue más tardía. Igualmente, diferenciar los grandes monos africanos (gorila y chimpancé) sólo a nivel de tribu, mientras el orangután se hace a nivel de subfamilia, implica una aceptación tácita de las modernas posturas acerca del significado evolutivo del tándem Sivapithecus-Ramapithecus, tal y como se verá a continuación. Debido a este estado de perpetua discusión, existen numerosos cuadros taxonómicos alternativos que reflejan diferentes posturas teóricas respecto a los diferentes problemas evolutivos que la cada vez más numerosa documentación va planteando.
Independientemente de este debate, el Orden de los Primates, en el que se incluye el género Homo, reúne a una serie de animales que presentan unas características generales bien marcadas, aunque siempre hay excepciones: (a) Buen desarrollo de la visión, fruto de una vida arborícola presente o pasada. Ojos en posición frontal, que proporcionan una visión en relieve y sensibilidad a una amplia gama de colores. (b) Manos y (la mayoría) pies prensiles, con pulgares oponibles y almohadillado en las plantas. (c) Uñas planas y normalmente cortas. (d) Sólo dos mamas en las hembras. (e) Dieta omnívora. (f) Alta sociabilidad. El orden de los primates se dividía tradicionalmente en antropoides (o monos verdaderos) y prosimios -arborícolas adaptados especialmente a la actividad nocturna-, entre los que se incluía el problemático Tarsio. Hoy en día se tiende a clasificar los primates en dos grandes grupos: Haplorrinos (con placa ósea tras las órbitas oculares), donde se incluyen todos los monos y los tarsiiformes, y Strepsirrinos (sin placa ósea), que corresponden a los antiguos prosimios. Dentro de los simios, la diferenciación principal se hace en función de la morfología nasal: Catarrinos (monos del Viejo Mundo) y Platirrinos (monos americanos). Los catarrinos siguen dividiéndose en dos superfamilias: Hominoidea (sin cola) y Cercopitecoidea (con cola, incluye a la mayor parte de los monos). Dentro de los hominoideos, las mayores novedades residen en la necesidad de señalar la mayor o menor proximidad genética (evolutiva) entre los cinco grupos principales (gibones, orangutanes, gorilas, chimpancés y hombres) determinada mediante pruebas bioquímicas (el denominado reloj bioquímico) que miden la cantidad de material genético común entre distintas especies; diferencias menores señalan fechas de divergencia específica más reciente.
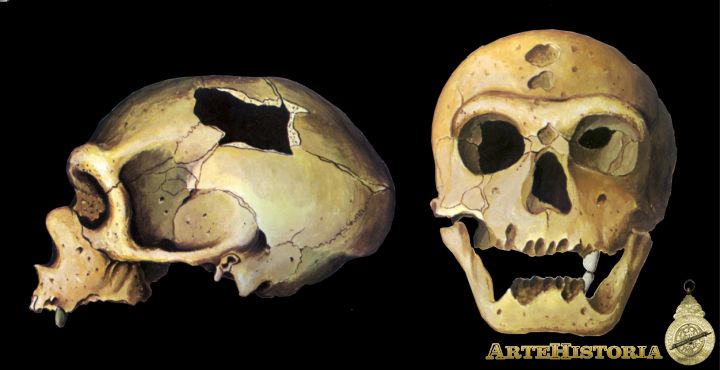
Este factor, junto a la paleontología, es el mayor determinante a la hora de reconstruir la historia de nuestro orden. Como se sabe, los mamíferos existieron bajo formas poco diversificadas durante todo el Mesozoico. Un fósil de estos mamíferos primitivos, denominado Purgatorius y datado a finales del Cretácico (c. 70 m.a.), se considera el primer representante del orden de los primates. Su morfología dentaria y su pequeño tamaño indican que se trataba de un insectívoro parecido a las musarañas actuales. A comienzos del Cenozoico los primates están representados por un nutrido grupo de animales arborícolas de pequeño tamaño (los plesiadápidos) de los que por especialización en los hábitos nocturnos se separaron los primeros strepsirrinos (Adapidos; c. 50 m. a.), antepasados de los lemures actuales. La evolución de los haplorrinos durante el Paleoceno y comienzos del Eoceno no es demasiado conocida, salvo en lo que hacía referencia al grupo de los Omomyiformes, antepasados comunes tanto de los tarsios como de los simios. A finales del Eoceno, con la apertura del Atlántico, se produce la definitiva separación de Catarrinos y Platirrinos, que quedan aislados geográficamente. La mejor documentación acerca de la diversificación de los catarrinos procede sobre todo de las asociaciones faunísticas de El Fayum (Egipto). Según las sucesiones de fósiles del Oligoceno, a partir del Oligopithecus y gracias a la expansión de los bosques se produce la separación de los cercopithecos (Paropithecus es su primer representante), mientras la línea evolutiva Propliopithecus-Aegiptopithecus se considera hoy en día el mejor candidato para el posterior desarrollo de los hominoideos.
A lo largo del Mioceno se produce la radiación de los hominoideos en tres grupos principales: los protogibones (Pliopithecus), los Proconsulidae, considerados hoy en día antepasados de los homínidos, y otros grupos extinguidos, como el famoso Oreopithecus bambolii, que también desarrollaron la braquiación como rasgo adaptativo a la vida arborícola. Entre los 15 y 10 m.a. se conoce una serie de hominoideos, descendiente de Proconsul, repartidos por Europa meridional, Asia y África oriental, en los que se han centrado numerosas controversias en los últimos años. Para algunos formaban un grupo homogéneo, los driopithecinos -cuyos restos también han aparecido en la Península-, mientras para otros se podían dividir en dos conjuntos: los driopithecinos s.s. y los ramapithecinos - grupo que englobaba a los fósiles de Ramapithecus, Sivapithecus y Kenyapithecus-, justificándose las diferencias de tamaño como dimorfismo sexual. Hoy en día, tras las oportunas rectificaciones de D. Pilbeam, se considera que Driopithecus y Ramapithecus son especies septentrionales, de Europa y Asia, sin descendencia evolutiva, mientras que Sivapithecus, simio de mayor tamaño circunscrito a Asia central y meridional, es el antepasado tanto de los orangutanes como del mayor primate que ha existido hasta ahora, el Gigantopithecus, que se extinguió al parecer ya en el Pleistoceno Medio. Por último, Kenvapithecus, descubierto por L. Leakey en Fort Ternan, es el mejor candidato para ser el antecesor común tanto de los homínidos como del gorila y el chimpancé. Entre los 10 y los 5 m.a. (Mioceno final) existe un enorme vacío de fósiles, achacable a los problemas de fosilización que presentan los primates por su tipo de vida, que impide precisar cómo se da el paso definitivo en la aparición de los homínidos. Los escasos restos datados a finales de este intervalo -un molar de Lukeino, el fragmento de mandíbula de Lothagam, el temporal de Chemeron y el húmero de Kanapoi, todos ellos procedentes de África Oriental-, son ya de homínido, aunque se discute sobre si alguno pertenece al género Homo o si todos son de Australopithecus arcaicos.
Independientemente de este debate, el Orden de los Primates, en el que se incluye el género Homo, reúne a una serie de animales que presentan unas características generales bien marcadas, aunque siempre hay excepciones: (a) Buen desarrollo de la visión, fruto de una vida arborícola presente o pasada. Ojos en posición frontal, que proporcionan una visión en relieve y sensibilidad a una amplia gama de colores. (b) Manos y (la mayoría) pies prensiles, con pulgares oponibles y almohadillado en las plantas. (c) Uñas planas y normalmente cortas. (d) Sólo dos mamas en las hembras. (e) Dieta omnívora. (f) Alta sociabilidad. El orden de los primates se dividía tradicionalmente en antropoides (o monos verdaderos) y prosimios -arborícolas adaptados especialmente a la actividad nocturna-, entre los que se incluía el problemático Tarsio. Hoy en día se tiende a clasificar los primates en dos grandes grupos: Haplorrinos (con placa ósea tras las órbitas oculares), donde se incluyen todos los monos y los tarsiiformes, y Strepsirrinos (sin placa ósea), que corresponden a los antiguos prosimios. Dentro de los simios, la diferenciación principal se hace en función de la morfología nasal: Catarrinos (monos del Viejo Mundo) y Platirrinos (monos americanos). Los catarrinos siguen dividiéndose en dos superfamilias: Hominoidea (sin cola) y Cercopitecoidea (con cola, incluye a la mayor parte de los monos). Dentro de los hominoideos, las mayores novedades residen en la necesidad de señalar la mayor o menor proximidad genética (evolutiva) entre los cinco grupos principales (gibones, orangutanes, gorilas, chimpancés y hombres) determinada mediante pruebas bioquímicas (el denominado reloj bioquímico) que miden la cantidad de material genético común entre distintas especies; diferencias menores señalan fechas de divergencia específica más reciente.
Este factor, junto a la paleontología, es el mayor determinante a la hora de reconstruir la historia de nuestro orden. Como se sabe, los mamíferos existieron bajo formas poco diversificadas durante todo el Mesozoico. Un fósil de estos mamíferos primitivos, denominado Purgatorius y datado a finales del Cretácico (c. 70 m.a.), se considera el primer representante del orden de los primates. Su morfología dentaria y su pequeño tamaño indican que se trataba de un insectívoro parecido a las musarañas actuales. A comienzos del Cenozoico los primates están representados por un nutrido grupo de animales arborícolas de pequeño tamaño (los plesiadápidos) de los que por especialización en los hábitos nocturnos se separaron los primeros strepsirrinos (Adapidos; c. 50 m. a.), antepasados de los lemures actuales. La evolución de los haplorrinos durante el Paleoceno y comienzos del Eoceno no es demasiado conocida, salvo en lo que hacía referencia al grupo de los Omomyiformes, antepasados comunes tanto de los tarsios como de los simios. A finales del Eoceno, con la apertura del Atlántico, se produce la definitiva separación de Catarrinos y Platirrinos, que quedan aislados geográficamente. La mejor documentación acerca de la diversificación de los catarrinos procede sobre todo de las asociaciones faunísticas de El Fayum (Egipto). Según las sucesiones de fósiles del Oligoceno, a partir del Oligopithecus y gracias a la expansión de los bosques se produce la separación de los cercopithecos (Paropithecus es su primer representante), mientras la línea evolutiva Propliopithecus-Aegiptopithecus se considera hoy en día el mejor candidato para el posterior desarrollo de los hominoideos.
A lo largo del Mioceno se produce la radiación de los hominoideos en tres grupos principales: los protogibones (Pliopithecus), los Proconsulidae, considerados hoy en día antepasados de los homínidos, y otros grupos extinguidos, como el famoso Oreopithecus bambolii, que también desarrollaron la braquiación como rasgo adaptativo a la vida arborícola. Entre los 15 y 10 m.a. se conoce una serie de hominoideos, descendiente de Proconsul, repartidos por Europa meridional, Asia y África oriental, en los que se han centrado numerosas controversias en los últimos años. Para algunos formaban un grupo homogéneo, los driopithecinos -cuyos restos también han aparecido en la Península-, mientras para otros se podían dividir en dos conjuntos: los driopithecinos s.s. y los ramapithecinos - grupo que englobaba a los fósiles de Ramapithecus, Sivapithecus y Kenyapithecus-, justificándose las diferencias de tamaño como dimorfismo sexual. Hoy en día, tras las oportunas rectificaciones de D. Pilbeam, se considera que Driopithecus y Ramapithecus son especies septentrionales, de Europa y Asia, sin descendencia evolutiva, mientras que Sivapithecus, simio de mayor tamaño circunscrito a Asia central y meridional, es el antepasado tanto de los orangutanes como del mayor primate que ha existido hasta ahora, el Gigantopithecus, que se extinguió al parecer ya en el Pleistoceno Medio. Por último, Kenvapithecus, descubierto por L. Leakey en Fort Ternan, es el mejor candidato para ser el antecesor común tanto de los homínidos como del gorila y el chimpancé. Entre los 10 y los 5 m.a. (Mioceno final) existe un enorme vacío de fósiles, achacable a los problemas de fosilización que presentan los primates por su tipo de vida, que impide precisar cómo se da el paso definitivo en la aparición de los homínidos. Los escasos restos datados a finales de este intervalo -un molar de Lukeino, el fragmento de mandíbula de Lothagam, el temporal de Chemeron y el húmero de Kanapoi, todos ellos procedentes de África Oriental-, son ya de homínido, aunque se discute sobre si alguno pertenece al género Homo o si todos son de Australopithecus arcaicos.